[論文発表]葉緑体の強光ストレス防御機構-APXとPGR5の相互関係@The Plant Journal誌
2021年5月18日
過剰な光エネルギーに対する植物の防御機構に関する丸田グループの論文がThe Plant Journal誌に受理されました。京都大学の山本先生との共同研究です。
Cooperation of chloroplast ascorbate peroxidases and proton gradient regulation 5 is critical for protecting Arabidopsis plants from photooxidative stress
Takashi Kameoka†, Takaya Okayasu†, Kana Kikuraku, Takahisa Ogawa, Yoshihiro Sawa, Hiroshi Yamamoto, Takahiro Ishikawa, Takanori Maruta*
The Plant Journal, 107: 876-892, 2021 Aug. DOI: https://doi.org/10.1111/tpj.15352
†equally contributed/ *corresponding author
[研究の概要]
光合成能力を上回る過剰な光エネルギーの存在下(強光ストレス)では、電子伝達系における活性酸素種(ROS、H2O2など)の生成が促進され、細胞が傷ついてしまいます。これを回避するために、植物は進化過程で葉緑体型アスコルビン酸ペルオキシダーゼ(ストロマ型sAPXおよびチラコイド膜結合型tAPX)を獲得し、生じたH2O2を速やかに消去することを可能にしました。さらに、チラコイド膜内腔の酸性化(∆pH形成)が「過剰光エネルギーの熱散逸」や「光化学系Iへの電子フラックスの抑制」などの仕組みを活性化することで、電子伝達の末端で生じるROS生成を抑制します。これらの仕組みには、∆pH形成に重要な役割を果たすProton gradient regulation 5(PGR5)タンパク質が関与しています。このように、光由来の酸化ストレスを防ぐためにさまざまな防御機構が存在しますが、それらの相互関係は不明でした。今回、私たちは葉緑体型APXとPGR5に注目し、全てを欠損させた三重変異株を用いた解析により、1)葉緑体型APXとPGR5は互いに相補的に機能しており、その協働が強光ストレス耐性に不可欠であること、2)pgr5変異株で見られる非光化学的消光(NPQ、熱散逸活性を含むパラメーター)の抑制が、三重変異株で少し回復すること、3)そして、その回復現象には循環的電子伝達経路(NDH経路)が関与することを見出しました。その他のさまざまなデータと合わせて、植物の光ストレスに対する極めて頑健な防御機構の概要を明らかにすることができました。
[研究の詳細]
1. 背景
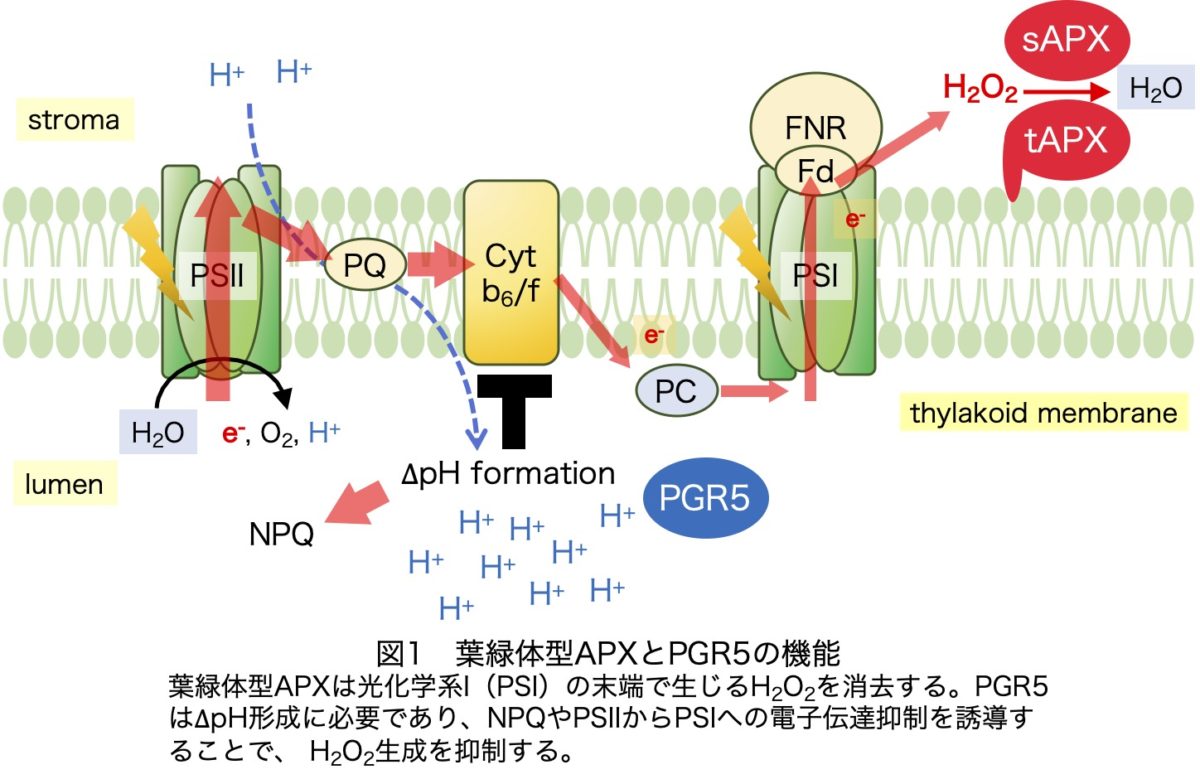
光合成を営む植物にとって、光はもっとも重要な環境要因です。しかし、光合成能力を上回る過剰光エネルギーの存在下では電子伝達系の過還元が起こり、その結果として活性酸素種(ROS、H2O2など)の生成が促進されます。適度な量であれば、H2O2はシグナルとして重要な機能を発揮しますが、過剰に蓄積すると細胞を傷つけてしまいます(光酸化的ストレス)。それを防ぐために葉緑体のストロマとチラコイド膜上にはアスコルビン酸ペルオキシダーゼ(それぞれsAPXおよびtAPX)が存在し、光合成で生じたH2O2を速やかに消去します(図1)。いくつかの説得力のある知見から、長らく葉緑体型APXは植物の強光ストレス耐性の鍵酵素であると信じられてきました。しかし、この考えとは対照的に、APXのノックアウト株は明確なストレス感受性を示しません。そのため、APXの抗酸化酵素としての機能や重要性は不明瞭でした(この一連の混乱はMaruta et al., Plant Cell Physiol., 2016に詳しくまとめています)。
抗酸化システム以外にも、光酸化的ストレスを防ぐ仕組みがたくさん存在します。特に、チラコイド膜内外の∆pHを介した防御機構が重要です。電子伝達に伴ってチラコイド膜内腔(ルーメン)が酸性化されると、キサントフィルサイクルが活性化され、過剰光エネルギーが熱として散逸されます。同時に、光化学系IIからIへの電子伝達が抑制されることで、H2O2(厳密にはスーパーオキシド)生成の場である光化学系Iの還元状態が防がれます。Proton gradient regulation 5(PGR5)は小さな葉緑体タンパク質であり、∆pH形成に不可欠な役割を担っています(図1)。そのため、pgr5変異株では光エネルギーの熱散逸や電子伝達抑制が起こりづらくなります。これらのPGR5依存的防御システムが光合成によるH2O2生成を抑制することで、葉緑体型APXの欠損を補う可能性が考えられますが、両システムの相互関係はほとんど不明でした。そこで今回、葉緑体型APXとPGR5の三重変異株を中心に、さまざまな多重変異株を作出して解析しました。
2. PGR5依存システムは葉緑体型APXを相補するか?
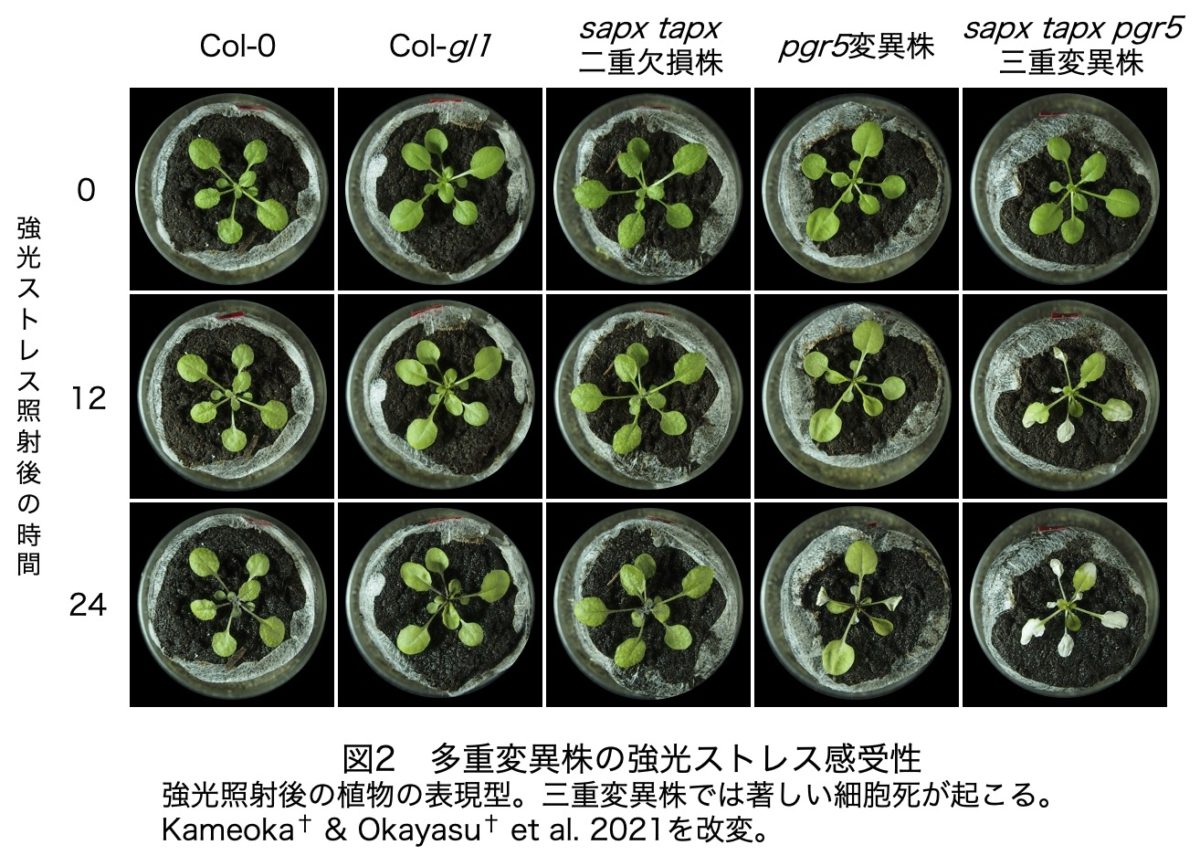
葉緑体型APXとPGR5依存システムの相互関係について調べるために、sapx pgr5およびtapx pgr5の二重変異株に加え、sapx tapx pgr5三重変異株を作出しました。そして、これらの株と、野生株および親株に強光ストレス(1,500 µmol photons/m2/s)を24時間照射しました。その結果、野生株やsapx tapx二重欠損株に視覚的なダメージは観察されませんでしたが、pgr5単独変異株では24時間後に部分的な葉の退色が見られました。この表現型は、sapx pgr5およびtapx pgr5の二重変異株とまったく変わりませんでしたが、三重変異株では著しく促進され、もっとも劇的な細胞死が見られました。これらの表現型の差は、光合成のダメージ度合いと綺麗に相関したことから、PGR5依存システムは葉緑体型APXを相補すること、そして逆も同じであることが明らかになりました。また、sAPXとtAPXはリダンダントな関係にあり、pgr5変異株で両方を欠損させた場合のみ劇的な細胞死が起こることがわかりました(図2)。
三重変異株でみられた強烈な細胞死がH2O2の蓄積および酸化損傷に起因するかどうか調べるために、H2O2に加えてアスコルビン酸およびグルタチオンを測定しました。その結果、いずれのデータも三重変異株における酸化ストレスの増大を支持しました。さらに、RNA-seq解析の結果から、三重変異株では酸化ストレス応答性遺伝子の発現が顕著に誘導されることがわかりました。これらの結果から、葉緑体型APXとPGR5依存システムは互いに相補的に機能すること、そして両者の協働は強光ストレス防御に不可欠であることが明らかになりました。なお、pgr5変異株では十分に∆pHが形成されないため、光ストレス防御に重要な「過剰光エネルギーの熱散逸」と「光化学系Iへの電子フラックスの抑制」の両方が制限されますが、後者が葉緑体型APXの相補に重要であることがさらなる遺伝学的解析により示唆されました。
3. pgr5変異株におけるNPQ誘導抑制がsapx tapx pgr5三重変異株では回復する?
pgr5変異株では過剰光エネルギーの熱散逸を十分に誘導することができないため、その指標となるクロロフィル蛍光パラメータNPQ(非光化学的消光)が低くなります。当然、野生株と比較するとsapx tapx pgr5三重変異株でもNPQ値は低くなったのですが、興味深いことに、pgr5単独変異株と比較すると高いNPQ値を示しました(図3)。また、キサントフィルの定量の結果から、pgr5におけるキサントフィルサイクル活性の低下が三重変異株では部分的に回復することもわかりました。
最近の研究により、光化学系Iの循環的電子伝達経路(NDH経路)がH2O2により活性化されることが示唆されています。NDH経路は光化学系Iの末端からプラストキノンへと電子を戻す仕組みで、∆pHの形成に寄与します。そのため、sapx tapx pgr5三重変異株では高蓄積したH2O2がNDH経路を活性化することで∆pHが少し回復し、その結果としてNPQも部分的に回復した可能性が考えられました。この可能性を検証するために、NDH経路を欠くcrr2変異株を用いて、sapx tapx pgr5 crr2四重変異株を作出しました。その結果として、コントロールとなるpgr5 crr2とsapx tapx pgr5 crr2四重変異株のNPQ値にまったく差がなく(図3)、sapx tapx pgr5三重変異株で見られるNPQ回復がNDH経路に依存的であることが強く示唆されました。

4. 結論
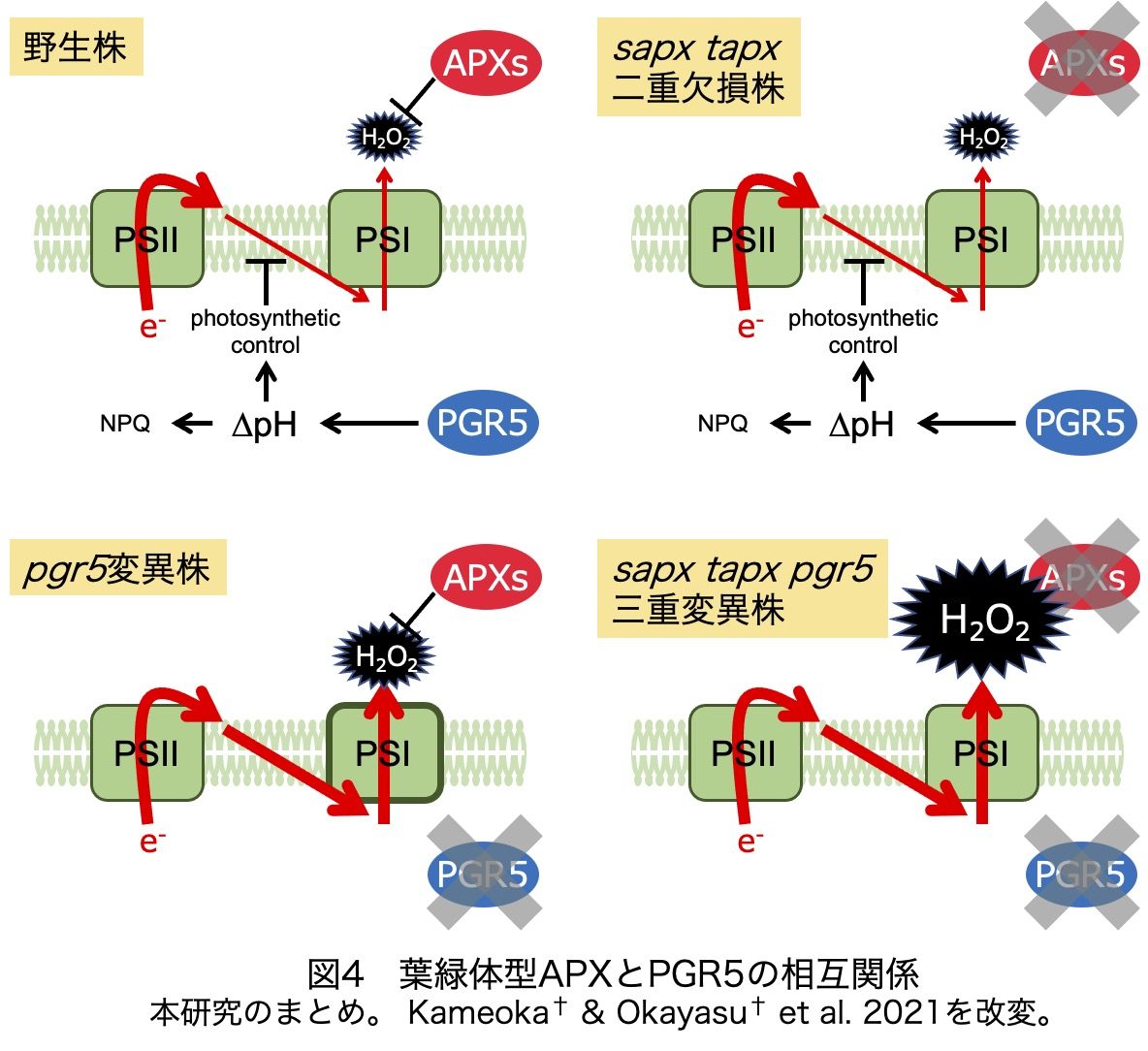
以上より、葉緑体型APXの機能はPGR5依存システムにより相補されること、その逆も然りであることが明らかになりました(図4)。つまり、葉緑体型APXの欠損株ではPGR5依存システムによりH2O2生成が強く制限されるために、強光ストレス条件で明確な表現型が現れません。一方、pgr5変異株ではH2O2生成速度が高まりますが、葉緑体型APXの働きにより、H2O2蓄積量が低く抑えられます。しかし、両方の仕組みを欠損すると、H2O2の蓄積が著しくなり、顕著な細胞死が起こります。興味深いことに、pgr5変異株で見られるNPQの誘導抑制が葉緑体型APXの欠損により少し回復し、この仕組みにはNDH依存の循環的電子伝達経路が関与するようです。おそらく、三重変異株では蓄積したH2O2がNDH経路を活性化し、∆pH依存の防御機構を部分的に回復させるのだろうと考えています。これは新しいストレス順応機構を意味するのかもしれませんが、とはいえsapx tapx pgr5三重変異株にとっては焼け石に水で、この三重変異株はボロボロになって枯れてしまいます。というワケで、本研究により、葉緑体型APXとPGR5依存システムの相互関係が明らかになり、植物は頑健な光酸化的ストレス防御機構を備えていることが明確になりました。
5. あとがき
本研究は修了生の岡安嵩也さんと亀岡峰志さんの修士論文研究で、同じく修了生(現・博士課程)の菊樂香奈さんにも手伝っていただきました。また、京都大学の山本先生にはDual-PAMを用いた光合成パラメータ解析をご指導いただきました。皆様、ありがとうございました。APXが「抗酸化酵素」として機能的かどうかについて、ずっと自信を持てずにいたので、pgr5バックグランドとはいえ、こうしてAPXが防御酵素として機能していることを自分たちで確認できて本当によかったです。思い返せば、岡安くんがsapx tapx pgr5三重変異株を作出したことが、丸田グループの怒涛の多重変異株作出ラッシュの引き金だった気がします。みんなたくさん多重変異株作りました(物凄い作業量)。でも、たまに多重すぎると否定されたりするんです(みんなゴメン)。それでも強調したいのは、植物は本当にたくさんの防御機構を冗長的に持っていて、一つ潰したくらいでは何も見えてこないということです。やっぱ多重じゃないと分からないことがたくさんあります。それは裏を返せば、植物の生きている自然環境がとても過酷であることを物語っています。きっと実験室では考えられないレベルのストレスを受けているはず!そんな視点の研究にも取り組み初めたりしています。それにしても、この論文は難産でした。審査過程では、こんなことあり得るのか!?と思うようなことも起こりました(初めてエディターとファイトしました)。数年したら良い経験だったと思えるのでしょう、メイビー(笑)