[論文発表]蘚類ヒメツリガネゴケにおけるアスコルビン酸生合成@The Plant Journal誌
2021年7月7日
蘚類ヒメツリガネゴケにおけるアスコルビン酸生合成の仕組みを明らかにした石川グループの論文がThe Plant Journal誌に受理されました。これは修了生の袖山 翼さん、西川 仁さん、原井健司さんらの研究をとりまとめたものです。
The D-mannose/L-galactose pathway is the dominant ascorbate biosynthetic route in the moss Physicomitrium patens
Tsubasa Sodeyama†, Hitoshi Nishikawa†, Kenji Harai†, Daiki Takeshima, Yoshihiro Sawa, Takanori Maruta, Takahiro Ishikawa*
The Plant Journal, 107: 1724-1738, 2021 Sep. DOI: https://doi.org/10.1111/tpj.15413
†equally contributed/ *corresponding author
[研究の概要]
高等植物において、D-マンノース/L-ガラクトース経路(スミルノフ経路)はアスコルビン酸生合成の主経路として機能します。また、その生理学的重要性は緑藻クラミドモナスでも報告されています。一方、陸上植物の基部にあたるコケ植物では報告例がありません。例えば、蘚類ヒメツリガネゴケにはスミルノフ経路の構成酵素遺伝子が全て揃っているため、本経路がドミナントに機能することが予想されるものの、実験的な裏付けがありませんでした。また興味深いことに、ガラクチュロン酸を介したオルタナティブ経路に関与する「アルドノラクトナーゼ(ALase)の相同遺伝子」がヒメツリガネゴケに存在することがわかり、コケ植物ではスミルノフ経路とガラクチュロン酸経路の両方が機能する可能性も示唆されました。そこで、生理生化学や分子遺伝学的手法を用いて解析した結果、1)ヒメツリガネゴケではスミルノフ経路がアスコルビン酸生合成に関与すること、2)ALaseは実際にALase活性を有しているものの、アスコルビン酸生合成には関与しないこと、3)この酵素は酸化型アスコルビン酸(デヒドロアスコルビン酸)の分解酵素として機能する可能性が示されました。これらの結果により、スミルノフ経路の普遍性に加え、コケ植物におけるアスコルビン酸の生理学的重要性が初めて明らかになりました。
[研究の詳細]
1. 背景
アスコルビン酸は優れた抗酸化剤であり、植物の主要レドックスバッファーです。高等植物において、アスコルビン酸はヘキソースリン酸プールからGDP-D-マンノースやL-ガラクトースを介して合成されます(図1左;スミルノフ経路)。これまでに単離されたシロイヌナズナのアスコルビン酸欠乏変異株(vtc1〜vtc4)の原因遺伝子のうち、VTC3以外の遺伝子はスミルノフ経路の構成酵素をコードすることから、この経路がアスコルビン酸生合成の主経路であることは遺伝学的に立証されています。VTC3タンパク質はキナーゼおよびホスファターゼの両ドメインを持つ風変わりなタンパク質で、スミルノフ経路の調節に関与すると考えられています。また、この経路は緑藻クラミドモナスにおいても必須であることから、緑藻から高等植物にいたる緑色植物において普遍的な経路であると考えられます。実際に、コケ植物、例えば蘚類ヒメツリガネゴケにおいてもスミルノフ経路の構成酵素遺伝子が全て保存されています。しかし、コケ植物における同経路の研究例はなく、アスコルビン酸生合成の進化や普遍性を議論する上での未解明の重要ポイントになっていました。

ところで、微細藻類ユーグレナはガラクチュロン酸を中間体とした経路(ガラクチュロン酸経路)でアスコルビン酸を合成します(図1右)。この経路においてL-ガラクトン酸からL-ガラクトノ-1,4-ラクトンへの変換(最終段階の一歩手前)を触媒するのがアルドノラクトナーゼ(ALase)で、この酵素は石川グループにより同定済みでした。ガラクチュロン酸経路は高等植物でも機能的である可能性が提唱されています。例えば、果実の成熟に伴うペクチンの分解によって大量のガラクチュロン酸が生成されるため、このような状況ではスミルノフ経路の代替経路として機能する可能性があります。しかし、高等植物にはALaseの相同遺伝子が存在しません。興味深いことに、ヒメツリガネゴケにはALaseの相同遺伝子が二つ存在(PpALase1および2)することがわかり、コケ植物ではスミルノフ経路とガラクチュロン酸経路の両方が機能的である可能性が示唆されました。そこで本研究では、ヒメツリガネゴケを実験材料として用いて、ALaseに加えて、スミルノフ経路の鍵酵素であるGDP-L-ガラクトースホスホリラーゼ(GGP/VTC2)およびVTC3の機能解析を試みました。
2. ヒメツリガネゴケにおけるアルドノラクトナーゼの存在
上述の通り、ヒメツリガネゴケには二つのALase相同遺伝子(PpALase1および2)が存在することがわかりました。PpALase1と2の同一性は16.7%と低いものの、類似性は50%程度で、両ALaseのアミノ酸配列はユーグレナALaseやヒトのオーソログ(SMP30)と30%前後の同一性を示しました。大腸菌を用いて両酵素のリコンビナント酵素を作製・精製し、酵素学的性質を調べたところ、PpALase1のガラクチュロン酸に対するKm値およびKcat値はそれぞれ2.98 mMおよび188.3 s-1でした。PpALase2も同程度のKm値を示したのですが、一方でKcat値は4.87 s-1と低く、その触媒効率(Kcat/Km)はPpALase1の40倍以上低いことがわかりました。同様の結果は、L-ガラクトノ-1,4-ラクトンを基質として用いた実験からも得られました。これらの結果から、PpALase1および2はALaseとしての酵素活性を有していること、特にPpALase1が主要酵素であることが強く示唆されました。
3. ヒメツリガネゴケはスミルノフ経路を使ってアスコルビン酸を合成する
次に、ALase1、あるいはスミルノフ経路がアスコルビン酸生合成に寄与するかどうかを調べる目的で、ALase1、VTC2およびVTC3の遺伝子破壊株を作出しました。ヒメツリガネゴケは三つのVTC2相同遺伝子(VTC2-1〜VTC2-3)を持っていましたが、これらの遺伝子の光応答性解析の結果、VTC2-3はほとんど光応答性を示さず、発現量も極めて低かったため、VTC2-1およびVTC2-2に焦点を絞りました。なお、ヒメツリガネゴケのVTC3遺伝子は一つでした。
野生株と比較して、VTC2-1およびVTC2-2破壊株の両方において著しいアスコルビン酸レベルの低下(それぞれ野生株の46%および17%)が認められました(図2)。この結果から、スミルノフ経路はヒメツリガネゴケにおいて機能的であることが初めて明らかになり、特にVTC2-2が重要であることがわかりました。

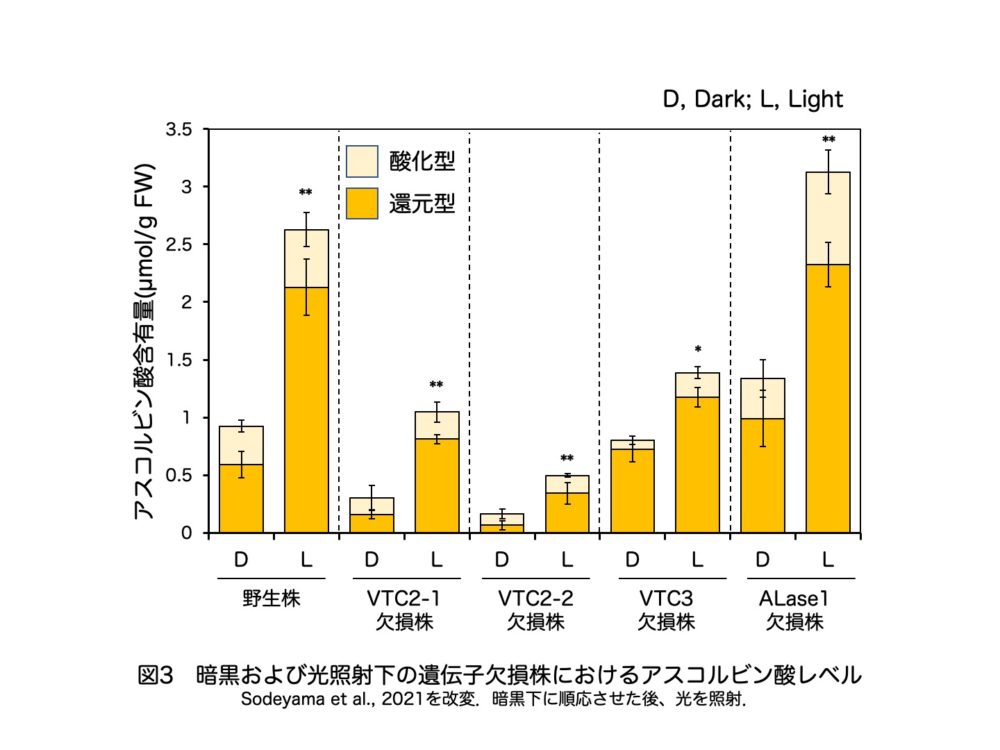
また、VTC3欠損株でもアスコルビン酸レベルの低下が生じました。さらに、この欠損株では暗順応後の光照射によるアスコルビン酸レベルの増加が顕著に抑制されました(図3)。これらの結果はシロイヌナズナの先行研究結果と一致しており、VTC3はコケ植物においてもアスコルビン酸生合成の光調節に関与することが明らかになりました。
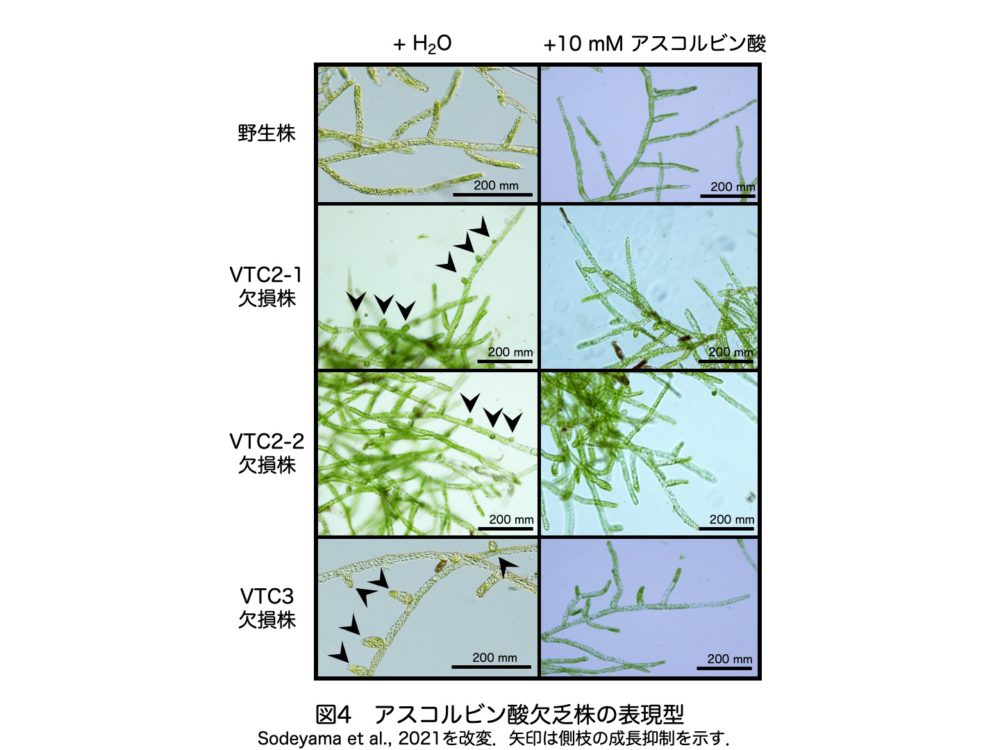
さらに、これらの欠損株では原系体の側枝の生育が抑えられ、この表現型は外部からのアスコルビン酸処理によって回復しました(図4)。したがって、スミルノフ経路を介したアスコルビン酸合成はヒメツリガネゴケの十分な成長に不可欠であることがわかりました。
一方、ALase1破壊株ではアスコルビン酸レベルの低下は起こらず、むしろ野生株と比較して有意に増加することがわかりました(図2)。この結果は、ALase1がアスコルビン酸生合成に関与しないことを意味しているのですが、興味深いことに、破壊株におけるアスコルビン酸の増加は還元型ではなく、酸化型(デヒドロアスコルビン酸、DHA)の増加に起因していました(図2右上)。さらに興味深いことに、2週齢までの時点では、ALase1破壊株は野生株と同様の表現型を示したのですが、3週齢以降に急速に茶褐色化し、細胞死が促進されたような表現型を示しました。このように、ALase1破壊株ではアスコルビン酸プールが増加しているにも関わらず、顕著な表現型を示すことがわかりました。
4. アルドノラクトナーゼはデヒドロアスコルビン酸の分解酵素?
なぜ、ALase1破壊株ではDHA量が増えるのでしょうか。過去にラット由来のラクトナーゼがDHAの分解反応を触媒することが報告されており、ALase1もDHA分解酵素として機能する可能性が考えられました。実際に、DHAに対する活性を調べたところ、VmaxおよびKm値はそれぞれ15.4 μmol min-1 mg-1 proteinおよび3.75 mMでした。ヒメツリガネゴケは高等植物の葉と同レベルのアスコルビン酸を含んでいます。そのうちの約10%程度が酸化型として存在するならば、ヒメツリガネゴケの細胞内にはmMオーダーのDHAが含まれると考えることができ、ALase1はDHA分解酵素として十分に機能的であると示唆されます。以上の結果を踏まえると、ALase1欠損株ではDHA分解反応が滞った結果としてDHAが蓄積したことが示唆されました。
5. まとめと考察
以上の結果より、高等植物や緑藻と同様に、ヒメツリガネゴケもスミルノフ経路を使ってアスコルビン酸を生合成することが明らかになり、本経路が緑色植物で高度に保存された主たるアスコルビン酸経路であることが明確になりました。紅藻もスミルノフ経路を利用すると考えられているのですが、面白いことにVTC2遺伝子を持っていません(他のスミルノフ経路の構成酵素遺伝子は全て揃っている)。そのため、VTC2と機能的に相同な別遺伝子が存在すると考えられていますが未解明です。スミルノフ経路は単細胞の緑藻で現在の形として確立された後、陸上植物で高度に保存されてきたのでしょう。ちなみに、灰色藻はスミルノフ経路を持たず、おそらくアスコルビン酸を合成しません。
もう一つの重要な発見は、高等植物におけるスミルノフ経路の調節因子・VTC3がヒメツリガネゴケでも機能的であったことです。VTC3は緑藻や紅藻にも存在するのですが(その機能は未解明)、緑藻の場合、スミルノフ経路は光よりも、むしろ活性酸素によって調節されているようですので、VTC3の機能は緑藻と陸上植物で多少異なる可能性があります。このあたりの調節因子の進化も今後の興味深い課題です。今、石川グループでは他のコケ植物にも興味を持って研究を進めていて、ヒメツリガネゴケと比較したときに、かなり興味深い特徴を持つことがわかってきました。なんだか面白い研究に発展しそうです。
さて、ヒメツリガネゴケにALase遺伝子が存在したことで、ようやくガラクチュロン酸経路が植物で機能的であることを証明できるかも!!と期待されたのですが、残念ながら(?)、少なくともヒメツリガネゴケにおいてALaseはアスコルビン酸生合成に関与しないようです。むしろ、この酵素は酸化型アスコルビン酸(DHA)の分解酵素として重要な役割を担う可能性が出てきました。DHAから還元型アスコルビン酸への再生にはグルタチオンが重要なのですが、ヒメツリガネゴケ(というかコケ全般?)のグルタチオン含量は極めて低く、アスコルビン酸再生能力も低いと思われます。DHAはタンパク質システインの酸化剤としても機能するため、潜在的に毒性です。DHAの毒性を回避するために、ヒメツリガネゴケはALaseを使って分解能力を高めているのかもしれません。それに対して、高等植物は高いアスコルビン酸再生能力を持っているので、DHA分解酵素としてのALaseが不必要なのかもしれませんね(実際に遺伝子が見つからない)。これらのストーリーが本当だったとしたら、ヒメツリガネゴケではアスコルビン酸のターンオーバーが速いはずです。それにも関わらず、高等植物と同レベルのアスコルビン酸を蓄積できるのはなぜなのか? そこまでしてアスコルビン酸を高蓄積する理由はなんなのか? まだまだ、興味深い謎が残されています。