[論文発表]グルタチオンを介したビタミンCの再生機構@Plant Physiology誌
2020年3月9日
強光ストレス条件で植物がビタミンCを高蓄積するためには、グルタチオンに依存した再生経路が極めて重要であることを丸田グループが明らかにしました。埼玉大学(川合先生、宮城先生)との共同研究であり、研究成果はPlant Physiology誌に受理されました。
Dehydroascorbate reductases and glutathione set a threshold for high light-induced ascorbate accumulation
Yusuke Terai, Hiromi Ueno, Takahisa Ogawa, Yoshihiro Sawa, Atsuko Miyagi, Maki Kawai-Yamada, Takahiro Ishikawa, and Takanori Maruta*
Plant Physiology, 183: 112-122, 2020 May. DOI: https://doi.org/10.1104/pp.19.01556
*corresponding author
さらに、本研究成果はPlant Physiology誌5月号にて、News and Viewsとして取り上げられました(↓)。
What Are the Roles for Dehydroascorbate Reductases and Glutathione in Sustaining Ascorbate Accumulation?
, Published May 2020. DOI: https://doi.org/10.1104/pp.20.00388
[研究の概要]
植物がビタミンC(アスコルビン酸)を還元型で高濃度に維持するためには、酸化型(デヒドロアスコルビン酸、DHA)からの再生系が重要です。今回、私たちはシロイヌナズナのデヒドロアスコルビン酸還元酵素(DHAR)の機能に注目し、様々な多重変異株を用いた解析から二つの結論を出しました。一つ目は、DHARの機能はアスコルビン酸濃度が高いときほど重要であり、強光条件下におけるアスコルビン酸のフル蓄積には必要であることです。一方で、アスコルビン酸濃度が低レベルまたは中程度のとき、DHARは重要ではなく、他の再生機構によって十分に相補されることが明らかになりました。二つ目は、DHARの機能を補う仕組みとして、グルタチオンによる非酵素的なDHA還元反応が重要であることです。DHARはグルタチオンを電子供与体としてDHAを還元する酵素であることから、グルタチオンを介した酵素的および非酵素的なDHA還元反応の協働が植物のアスコルビン酸再生に相補的に機能することが明らかになりました。そして三つ目は、この協働作業がアスコルビン酸の高蓄積および強光ストレス耐性に不可欠であることです。Open Accessなので、興味あれば是非読んでください。
[研究の詳細]
1. 背景
アスコルビン酸は植物にもっとも豊富に存在する水溶性抗酸化剤であり、細胞内のレドックス恒常性の維持に不可欠です。アスコルビン酸は活性酸素種を消去すると、自身は一電子酸化されモノデヒドロアスコルビン酸(MDHA)ラジカルとなります。このラジカルは続く自発的な不均化反応によって、アスコルビン酸と二電子酸化型のデヒドロアスコルビン酸(DHA)となります。DHAは極めて不安定であるため、アスコルビン酸を還元型として高濃度に蓄積するためには、酸化型からの再生が不可欠です。
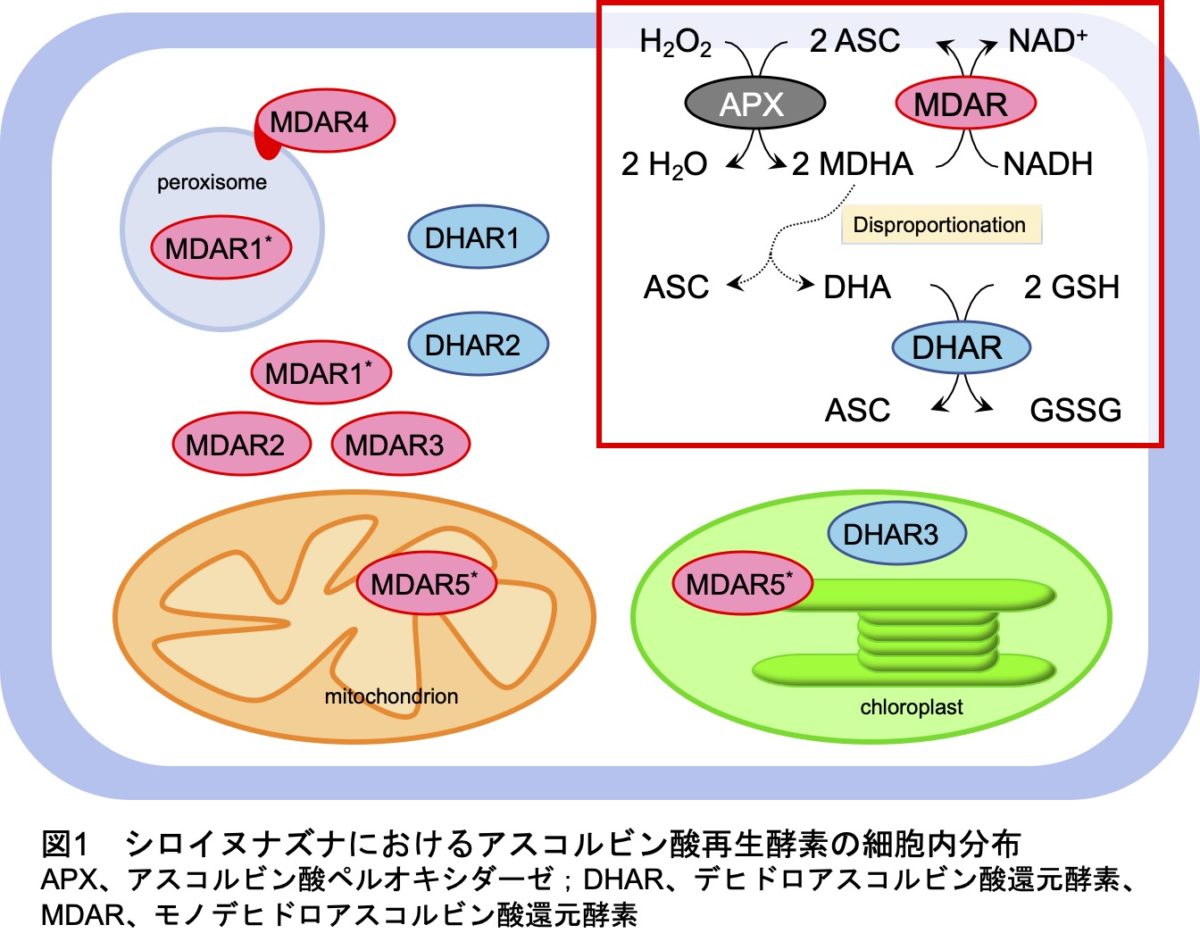
植物はアスコルビン酸再生酵素としてモノデヒドロアスコルビン酸還元酵素(MDAR)とデヒドロアスコルビン酸還元酵素(DHAR)を持っています。モデル植物であるシロイヌナズナには、それぞれ5および3のMDARおよびDHARアイソフォームが存在し、図1に示す細胞内局在性を示します。これらの酵素の協調的なはたらきが還元型アスコルビン酸の維持に重要だと考えられていますが、遺伝学的な見地からの証明はありませんでした。そこで今回、私たちはDHARの生理学的重要性について、特に強光ストレス下のアスコルビン酸再生に着目し、シロイヌナズナの遺伝子破壊株を用いて検証しました。
2. 強光ストレス下におけるアスコルビン酸高蓄積へのDHARの関与
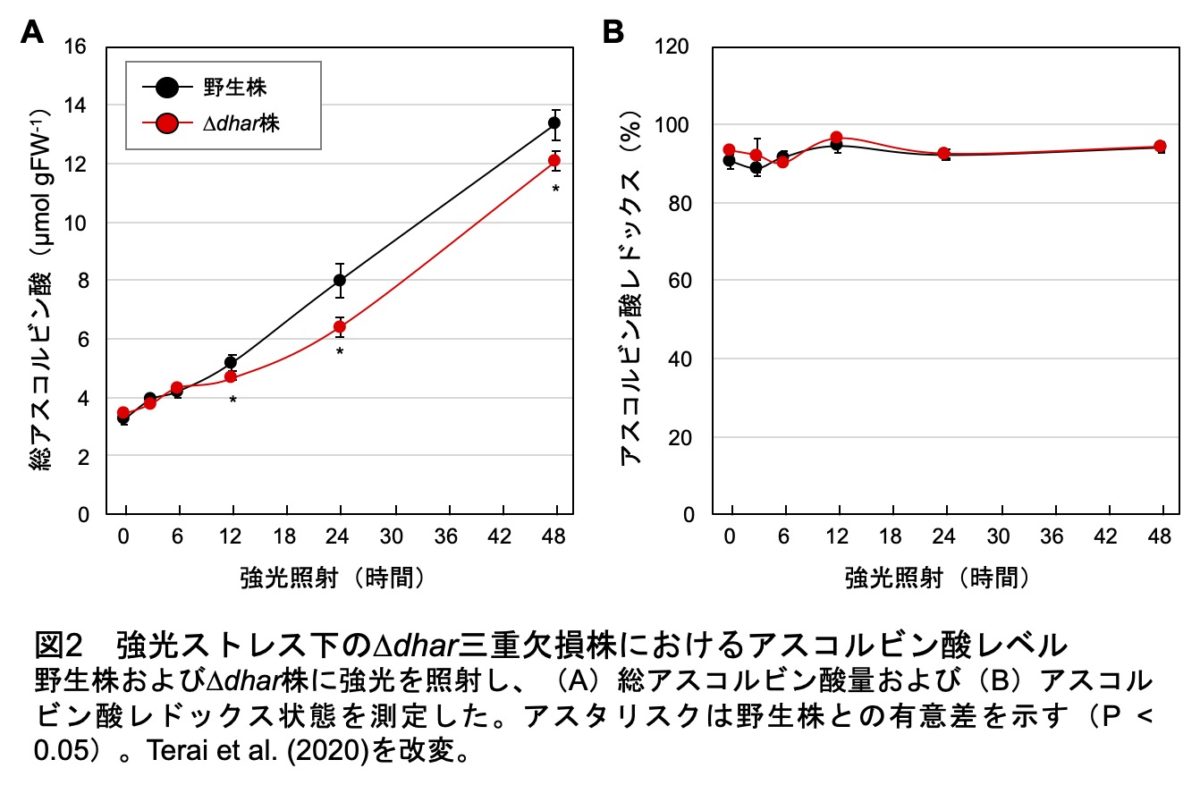
まず、シロイヌナズナのDHAR1、2および3の欠損株を単離しました。さらに、それぞれを掛け合わせて二重欠損株、さらには三重欠損株(便宜上、∆dharと呼びます)まで作出しました。結果として、∆dhar株ではDHAR活性がほぼゼロまで低下したのですが、アスコルビン酸の量やレドックス状態に野生株との差は見られませんでした。そこで、アスコルビン酸の生合成とターンオーバーを活性化するために、野生株と∆dhar株に強光ストレス(1,500 µmol photons/m2/s)を付与しました。その結果、野生株では強光照射に伴って著しくアスコルビン酸レベルが増加し、この増加は∆dhar株で統計学的には有意に抑制されました(図2)。しかし、その差はごくわずかであり、∆dhar株は野生株レベルのストレス耐性能を保持しました。これらの結果から、DHARが重要かどうかますます不明瞭になってしまいました。
3. アスコルビン酸濃度とDHARの機能の関係
野生株と∆dhar株の差はアスコルビン酸レベルが高くなる強光照射12時間後以降でのみ生じたことから、DHARの重要性はアスコルビン酸濃度依存的であり、アスコルビン酸濃度が高いときほど必要性が増す可能性が示唆されました。このことを詳細に検討するために、アスコルビン酸生合成変異株であるvtc2を用いました。vtc2株では野生株の約30%までアスコルビン酸が低下します。この株を∆dhar株と掛け合わせ、∆dhar vtc2四重欠損株を作出しました。結果として、強光照射下の∆dhar株および∆dhar vtc2株のアスコルビン酸レベルに有意な差は認められませんでした。つまり、野生株と∆dhar株を比較した場合に見られるアスコルビン酸レベルの差が、vtc2バックグラウンドでは消失するということです。したがって、DHARはアスコルビン酸濃度が高いときほど重要であることが強く示されました。おそらく、低濃度のときはアスコルビン酸の酸化速度が低下し、他の再生系で十分に賄えるのだと考えられます。
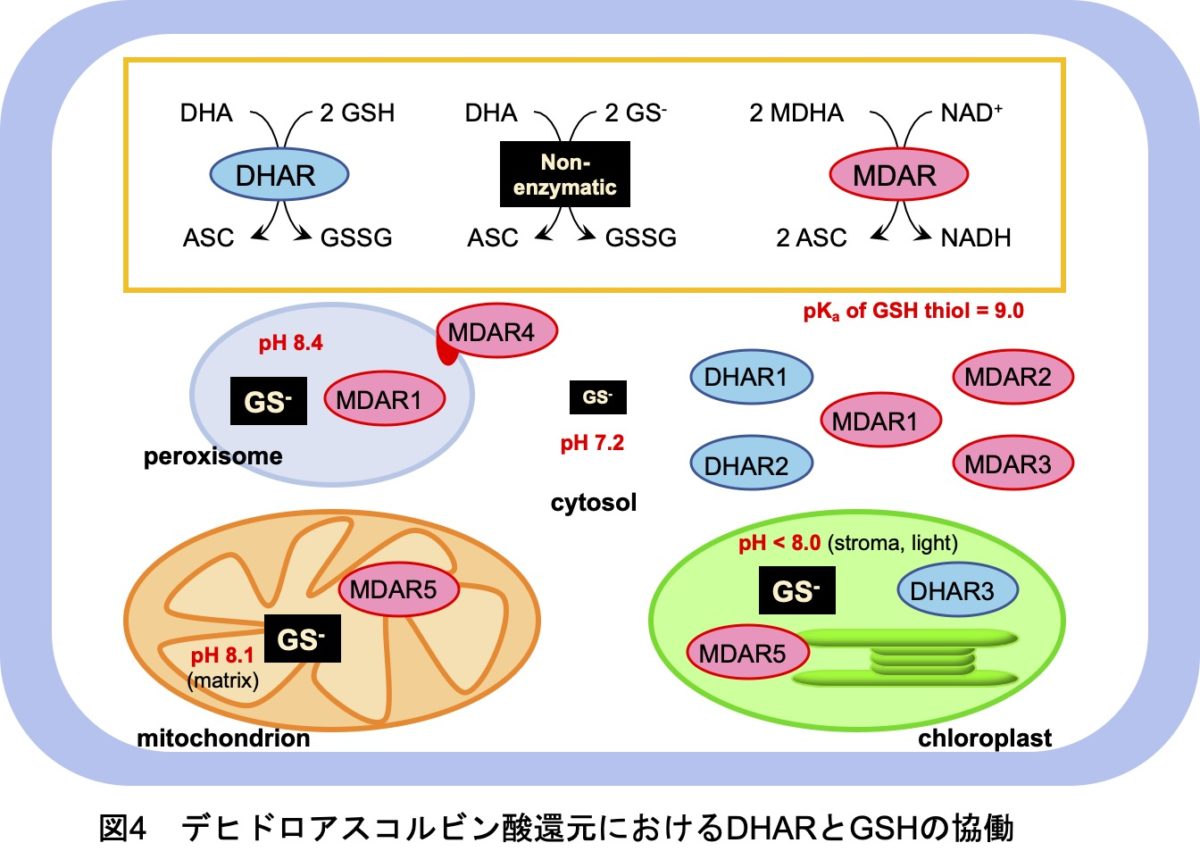
4. 還元型グルタチオンはDHARの機能を相補する
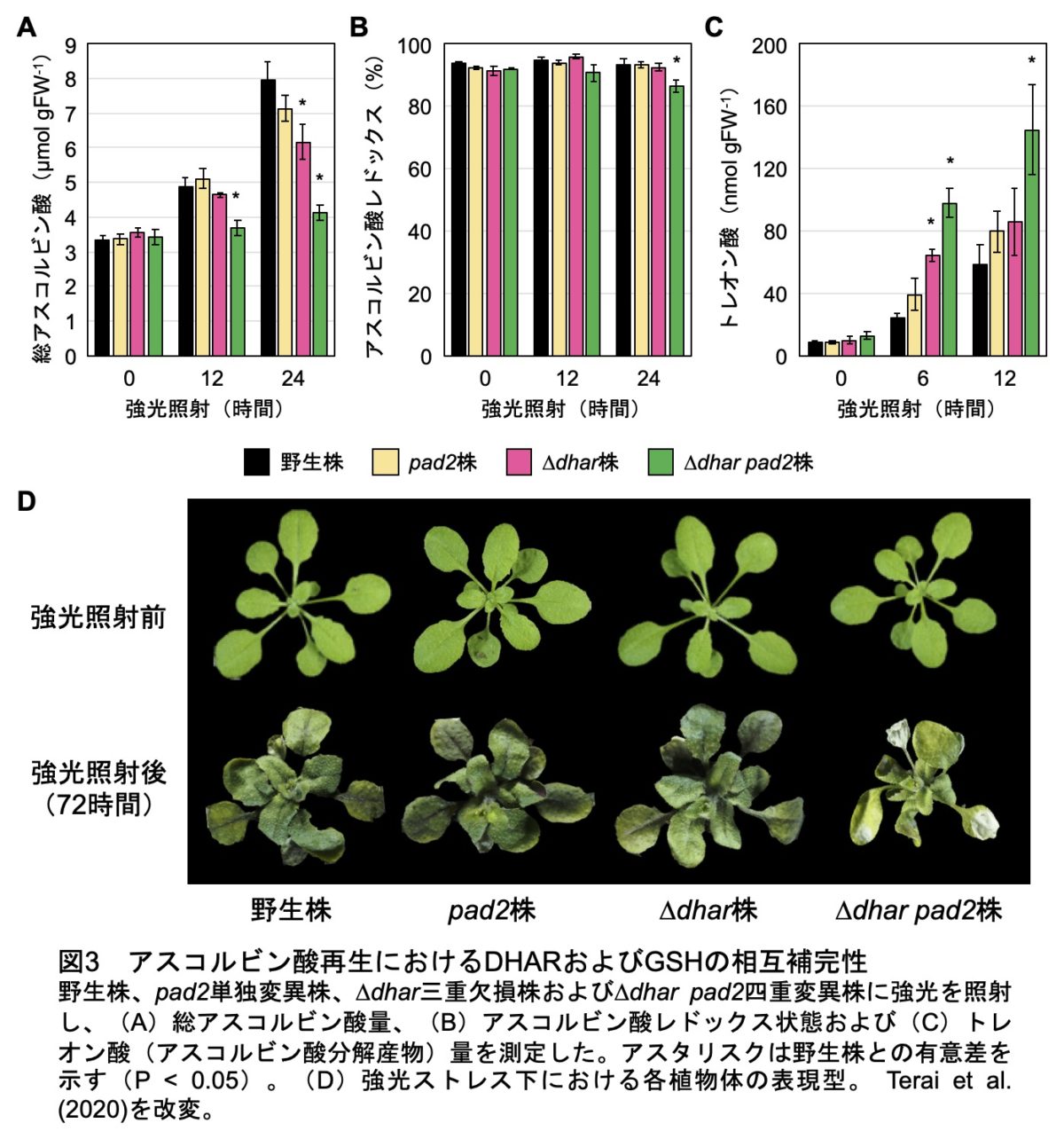
次に、DHARを補う別のDHA還元系として、私たちは還元型グルタチオン(GSH)そのものに注目しました。GSHはDHARの電子供与体ですが、自身も非酵素的な反応によりDHAを還元することができます。ただ、この非酵素反応はGSHの脱プロトン化(GS-アニオン)を必要とし、GSHチオールのpKaは約9.0と高いため、生理学的な条件でどれほど有効かは未解明でした。GSHによる相補の可能性を検証するために、GSH欠乏変異株(pad2)を用いました。この株は野生株の約30%のGSHを保持します。DHARのGSHに対するKm値がmMオーダーと高いことを考慮すると、この変異株ではGSHによるDHA還元に加えて、DHAR反応も抑制されると期待されました。しかし予想とは裏腹に、強光ストレス条件においてpad2変異株は野生株レベルでアスコルビン酸を蓄積することが明らかになりました。
さらなる検証のため、pad2と∆dharを交配し、∆dhar pad2四重変異株を作出しました。その結果、∆dhar pad2四重変異株では強光照射によるアスコルビン酸レベルの増加がほぼ完全に抑制されました(図3)。また、∆dhar株へのGSH生合成阻害剤処理の実験からも、同様の結果が得られました。一方、アスコルビン酸のレドックス状態への影響はわずかでした。この原因は、DHAの不安定生に起因すると考えられました。そこで、アスコルビン酸のターンオーバーを調べるために、分解産物量を定量したところ、強光条件の∆dhar pad2四重変異株において高レベルでの蓄積が認められました(図3)。したがって、∆dhar pad2四重変異株ではアスコルビン酸のターンオーバーが高まっていることが明らかになり、レドックス状態に影響が出なかったのはDHAが速やかに分解されるためであると考えられました。さらに、∆dhar pad2四重変異株は強光ストレスに対して著しい感受性を示したことから(図3)、DHARとグルタチオンの協働によるアスコルビン酸再生は、強光ストレス耐性に不可欠であることも証明できました(図4)。
5. あとがき
本研究は、修了生の寺井くんと始めたものです。実は当初、DHARのアスコルビン酸再生酵素としてのはたらきより、むしろ「GSH酸化酵素」の側面に注目して研究を開始しました。酸化ストレス条件でのグルタチオン酸化およびレドックス変化にDHARが関与するかどうかが知りたかったのです。しかし直後に、強力なコンペティターが強烈なデータを出していることを偶然知り、そのテーマでは彼らに追いつけないことに気づきました。慌てて、DHA還元反応(すなわち従来のDHARの役割)に視点を変えて研究することにしましたが、予期せぬ結果の連続(しかも、僕たちを狂喜乱舞させた一発限りの再現不可現象もあった笑)で、DHARは本当にアスコルビン酸再生酵素なのか?と何度も疑いながら、何度も自分たちの頭の中のストーリーを書き換えながら進めました。
今回の報告のキモは、∆dhar pad2四重変異株です。これは、DHAR酵素反応に加えて、GSHによる非酵素反応も抑えるために作ったものですが、そもそもDHARはGSH依存酵素(しかもGSHへのKm値は比較的高い)なので、pad2のシングル変異株でも酵素/非酵素反応の両方が抑えられると思われました。そのため丸田は、四重変異株を作る価値は低いと思う旨を寺井くんに伝えていたのですが、彼は「せっかくなんで」とそのまま作出を継続しました。その結果として、四重変異株でのみ著しいアスコルビン酸ターンオーバーの促進と葉のブリーチングが起こり、今回の発見に繋がりました。おそらく彼がいなければDHARの生理機能やインパクトは未だに分からないままで、DHARに「役立たずな酵素」という誤ったレッテルを貼っていたかもしれません。寺井くんには感謝感激雨霰です。また、そのうち浴びるほど一緒に酒を飲みたいですね。
寺井くんの修了後、卒業生の上野さんがバトンを引きつぎ、おそるべき精度で必要なデータを積み重ねてくれました。また埼玉大学の宮城先生には、アスコルビン酸分解産物の定量を引き受けていただきました。みなさま、ご協力ありがとうございました。